|
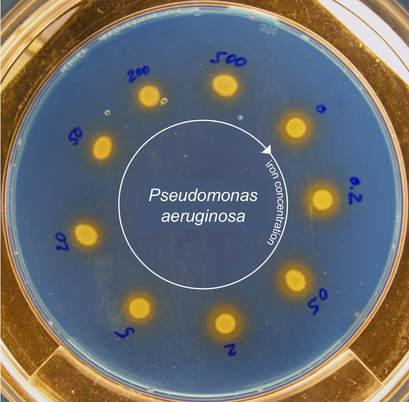
Diffusible public goodsWe are interested in beneficial compounds secreted by bacteria, which can be shared as public goods between cells. Iron-chelating siderophores are examples of such public goods. The diffusible nature of these molecules becomes visible on CAS-plates, where siderophores induce a colour change from blue to orange when chelating iron outside the cells (orange halos around yellow colonies). Siderophores are costly to produce and their production is fine-tuned in response to iron availability. This is illustrated by the halos becoming smaller when bacteria were cultured with increased levels of iron (blue numbers in µM) prior to spotting onto the plate. |
Cooperation and cheatingDuring experimental evolution, we repeatedly observe the spread of mutants that stop producing public goods, even under conditions where they are actually required for growth. In the case of pyoverdine (the green-fluorescent siderophore produced by Pseudomonas aeruginosa), the spreading of non-producers can be spotted by the naked eye, as cultures become less green (see image). We are interested in understanding whether these non-producers evolve as cheaters, exploiting the public goods produced by the wildtype, and what factors can prevent the collapse of cooperation. |

|
|
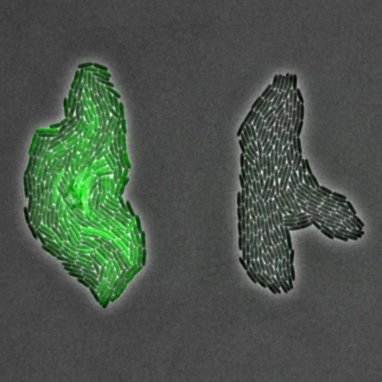
Bacterial interactions at the cell levelWe explore the social interaction patterns between bacteria at the single-cell level. The picture shows a colony of wildtype bacteria of P. aeruginosa on the left, which highly invests into pyoverdine production (green fluorescent protein expression fused to the pyoverdine synthesis gene pvdA). The wildtype grows next to a colony of pyoverdine non-producers. Using this single-cell approach, we can follow colonies over time, and ask whether producers sense the presence of exploitative non-producers and adjust gene expression accordingly, and whether the relative success of non-producers depends on the proximity to the producers. |
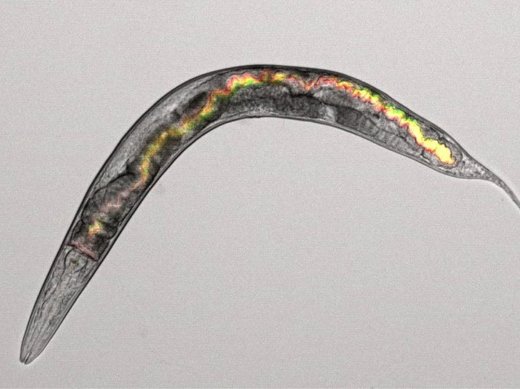
Public goods as virulence factorsWe study secreted public goods in the context of infections. For this purpose, we use the nematode Caenorhabditis elegans as a host model system. Worms are transparent, which enables us to monitor infections inside hosts. The gut of the individual shown in the picture, for instance, is colonized by two strains of P. aeruginosa, each labelled with a constitutively expressed fluorescent marker (eGFP and mCherry, overlay in yellow). Using this approach, we can study: (i) the expression of virulence factors inside hosts (via gene-reporter fusions), (ii) strain interactions, and (iii) the damage inflicted on the host. Since C. elegans naturally grazes on bacteria, we can also setup microcosms with hosts and bacteria, and study how bacterial virulence evolves over time. |
|
|
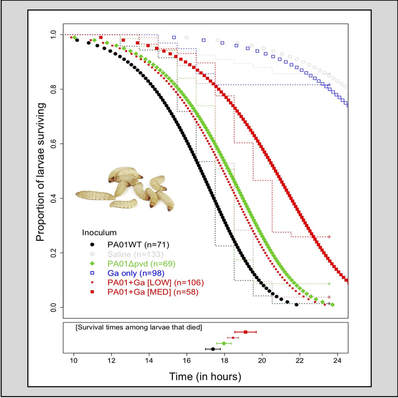
Infections and public good quenchingSince secreted bacterial compounds are often virulence factors inside hosts, there is great interest in therapeutic approaches that interfere with virulence factor production. We are interested in approaches that disable secreted siderophores from functioning, thereby inducing iron starvation and reducing pathogen growth. One such approach consists in the addition of gallium to infections, a metal ion that irreversible binds to pyoverdine produced by P. aeruginosa. In infection experiments using the larvae of the greater wax moth (Galleria mellonella), we found that gallium treatment (red curves) significantly enhances host survival compared to untreated larvae (black curve) because it disables pyoverdine-mediated iron acquisition. |
|
Social strategies & community dynamics We are interested in understanding how different types of social interactions affect the composition and stability of multi-strain communities. We study this in the context of pyoverdine production, a siderophore for which different chemical variants exist, each relying on a specific receptor for uptake. One of our findings is that the presence of multiple social strategies can stabilise community diversity. This is shown in the triangle plot, where three strains chase each other in circles with no overall winner. C produces a potent pyoverdine type, and thus outcompetes L producing a less efficient pyoverdine type. However, C is vulnerable to exploitation by D, while L is resistant to cheating because it produces a non-exploitable unique pyoverdine type. |

|
|
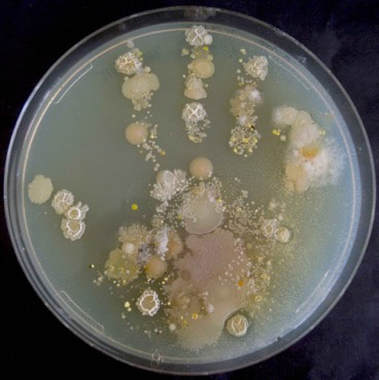
Social interactions in natural communities Natural microbial communities are extremely diverse. This is nicely illustrated by the depicted handprint of a child on an agar plate. In our studies, we focus on the extremely diverse clade of Pseudomonas bacteria. We have assembled a strain collection of several hundred strains, from different soil and freshwater communities. We have quantified the social profile of each strain - its ability to make siderophores, proteases, biosurfactants, biofilms and toxins. We now reassemble these communities in the lab and examine how the different types of social interactions shape community diversity and stability. |
